Эволюция древнейших предков однокопытных в третичном периоде
Современные лошади и родственные им формы однокопытных животных относятся к классу млекопитающих, сверхотряду копытных (Ungulata), отряду непарнокопытных (Perissodactyla), семейству лошадей (Equidae), роду лошадей (Equus). Род лошадей подразделяется в свою очередь на четыре подрода:
1) подрод собственно лошадей (Equus), к которому относятся современные виды домашних лошадей, вид дикой лошади Пржевальского (Е. Przewalskii Р.), а также ранее существовавший тарпан (Е. с. Cmelini);
2) подрод зебр (Е. Hippotigris), с видами: Н. zebra, H. Quagga, Н. grevii и др.;
3) подрод ослов (Asinus) с современными видами африканских ослов, распространенных в Африке, Азии и других странах света.
Отдельные ученые (В. Громова) выделяют азиатских полуослов из подрода ослов (Asinus) в самостоятельный четвертый подрод (Е. Hemionus), к которому относят современные дикие формы: куланов, онагров, киангов. Это подтвердили и опыты по гибридизации, проведенные В. А. Щекиным и А. В. Шкургиным (1933-1950 гг.).
В выяснении истории эволюции копытных животных важнейшую роль сыграли работы выдающегося русского ученого В. О. Ковалевского (1842-1883). Его монографии об анхитерии (древний представитель непарнокопытных из нижнего миоцена), о хиопотамусе (Hyopotamus, из древних парнокопытных) и другие работы имели большое значение для развития эволюционной теории и нанесли сокрушительный удар идеалистическим взглядам в палеонтологии о неизменяемости видов
В. О. Ковалевский является основоположником современной эволюционной палеонтологии. В своих работах об анхитерии (1873 г.) он доказал существование переходных форм ископаемых однокопытных животных, что отрицал Кювье, и сформулировал основные закономерности эволюции этой группы животных.
Анхитерий, по Ковалевскому, - это один из промежуточных родов в генеалогической линии непарнокопытных. Ковалевский писал о нем следующее: "... анхитерий по строению своего скелета является столь промежуточным, переходным родом, что если бы теория трансмутации не была уже прочно обоснована, он мог быть одной из наиболее важных ее опор"*.
* (В. О.Ковалевский, Палеонтология лошадей, изд. Академии наук СССР, 1948, стр. 15.)
Работы В. О. Ковалевского не только выясняют направление и основные этапы эволюции предков копытных животных, но и устанавливают, что эта эволюция шла под влиянием изменившихся условий существования - постепенного перехода от лесного образа жизни к жизни на открытых пространствах.
Приспособление к изменявшимся условиям вызывало постепенное изменение функции и строения животных, особенно ярко обнаруживающееся в строении конечностей и зубного аппарата.
Жизнь на открытых пространствах и более плотных почвах развивала способность к быстрому бегу и вызывала редукцию (утрату) боковых пальцев, а переход на более жесткую травянистую пищу обусловил изменение и усложнение зубного аппарата.
Предки лошадей встречаются как в Европе, так и в Северной Америке начиная еще с древнейших слоев третичного периода. В связи с тем, что Северная Америка была связана с Азией сушей и многие ископаемые формы общи для обеих частей света эволюцию лошадей в третичный период большинство советских ученых рассматривает в общем плане, включая в нее как формы, обнаруженные в Европе, так и промежуточные формы, найденные в Северной Америке.
Заключение ряда ученых о том, что основная линия эволюции лошадей в третичный период шла в Северной Америке (где известно много промежуточных форм), а в Евразию мигрировали из Америки и расселялись только отдельные роды, вызывает возражения, так как третичные отложения в Азии и Европе пока недостаточно изучены; в дальнейшем здесь могут быть найдены недостающие промежуточные звенья.
Древнейшие предки лошади были лесными животными, обитавшими во влажных тропических лесах, распространенных в первую половину третичного периода (в эоцене и олигоцене)* в умеренных широтах Старого и Нового Света. Они были приспособлены к жизни в лесу на влажных и рыхлых почвах и питались сочной лесной растительностью. Эволюция древнейших представителей предков лошади шла в этот период в направлении укрупнения размеров, усложнения зубного аппарата и развития способности к быстрому передвижению по рыхлому и вязкому лесному грунту. Одновременно с удлинением конечностей шло их приспособление к прямолинейному движению, сопровождавшееся укреплением блоковидных суставов и частичной редукцией боковых пальцев при сохранении опоры на три пальца. При этом средний палец приобретал большие размеры и нес основную нагрузку, боковые же пальцы уменьшались и укорачивались, сохраняя роль дополнительной опоры, позволяющей передвигаться по рыхлой почве.
* (Третичный период - это первый, самый древний период последней - кайнозойской - эры (эры млекопитающих и человека). Он подразделяется на пять эпох: палеоцен (самая древняя, начавшаяся более 50 млн. лет назад), эоцен, олигоцен, миоцен и плиоцен. За плиоценом наступил четвертичный период.)
Древнейшие предки непарнокопытных животных обнаружены в нижних (самых древних) отложениях третичного периода Европы и Северной Америки. Эти животные, принадлежавшие к роду фенакодов (Phenacodus), имели пятипалые конечности, опирались на три средних растопыренных пальца, из которых наиболее развитым был средний, одетый широким копытцем. В переднем поясе ключицы уже не было, локтевой сустав был выпрямлен, как у высших однокопытных (это указывает на утрату хватательной функции и приспособление к быстрому передвижению). Бугорчатое строение зубов указывало на приспособленность к мягкой, сочной растительной пище.
Один из видов рода Phenacodus - tetraclaenodon puercensis S. (из нижнего эоцена), выделяемый отдельными учеными в самостоятельный род, ставится у основания семейства лошадиных*.
* (См. проф. М. В. Павлова, Палезоология, ч. II, 1929, стр. 233.)
Следующей ступенью в эволюции непарнокопытных животных являются хиракотериум (Hyracotherium), обнаруженный в нижнем и среднем эоцене Европы и эогиппус, обитавший в тот же период в Северной Америке.
Хиракотериум - это еще мелкое (30-50 см) животное с более тонкими и более длинными, чем у фенакода, конечностями, из которых передние имеют по четыре пальца, а задние - по три. Следы первого пальца на передних ногах исчезли, на задних ногах сохранились рудименты двух крайних пальцев в виде грифельных косточек. Голова у хиракотериума относительно более крупная, чем у фенакода, с вытянутой и узкой мордой и несколько более сложными, но еще типично бугорчатыми зубами*.
* (См. А. Борисяк, Курс палеонтологии, ч. II, 1906, стр. 297.)
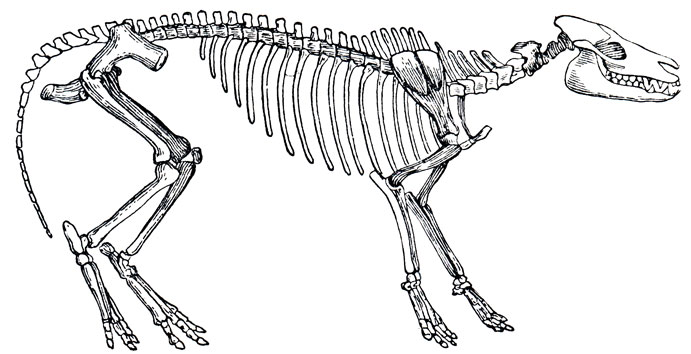
Рис. 83. Скелет хиракотериума (Hyracotherium ventjculum cope)
Эогиппус (Eohippus) имел конечности с четырьмя пальцами на передних и тремя на задних ногах и несколько более осложненные зубы.
В олигоцене жила форма древних непарнокопытных, характеризовавшихся дальнейшей редукцией пальцев конечностей и некоторым усложнением зубного аппарата. Это был Мезогиппус (Mesohippus), размерами с мелкую овчарку, и более крупный миогиппус. Судя по строению зубов, они питались мягкой растительностью в лесах; имели трехпалые конечности с более длинными метаподиями; раздвинутые пальцы позволяли передвигаться по рыхлой почве. От четвертого наружного пальца передней ноги остался рудимент его пястной кости; средний палец по размерам был несколько крупнее остальных, но все три несомненно достигали земли. Хвост укоротился. Зубы были более сложные и более прочные, чем у Hyracotherium. Усложнение зубного аппарата этой формы проявляется в образовании гребней и лунок на поверхности коренных зубов, предкоренные зубы по своему строению приближаются к коренным*.
* (См. А. Ш. Ромер, Палеонтология позвоночных, перевод под ред. Л. Ш. Давиташвили. ГОНТИ, 1939.)
В конце олигоцена и начале миоцена в Европе и Америке получил распространение анхитерий (Anchitherium), детально изученный В. О. Ковалевским. Это довольно крупное животное, достигавшее размеров пони, имело трехпалые конечности с более массивным и более длинным средним пальцем; крайние пальцы были короче и тоньше среднего, но все же достигали земли. Зубы анхитерия хотя и были несколько сложнее, чем у предшествовавших форм, но оставались еще низкокоронковыми, не имели постоянного роста, что указывало на употребление мягкой пищи и жизнь в лесах. По мнению ряда ученых, анхитерий не находится в прямом ряду предков лошади.
Изучая зубной аппарат ископаемых форм копытных животных, В. О. Ковалевский установил, что в середине миоцена произошло коренное изменение в строении этого аппарата, что могло быть вызвано только переходом копытных на новую, более жесткую пищу. Это заключение блестяще подтвердилось работами ботаников, установивших в миоцене неизвестную ранее травянистую растительность*.
* (А. А. Борисяк, В. О. Ковалевский, его жизнь и научные труды, 1928, стр. 110.)
Постепенное похолодание и уменьшение влажности климата влекли за собой изменение растительности и условий существования животных. Тропические леса в Европе к концу третичного периода оттесняются из средней полосы на дальний юг. На их месте развиваются в северной полосе хвойные и смешанные леса, а к югу от этих лесов - жаркие саванны и сухие степи. В миоцене на обширных равнинах получает пышное развитие травянистая растительность, которая становится пищей и важнейшим условием процветания травоядных животных, заселявших равнины.
Переход к жизни на открытых пространствах, с заменой мягкой и сочной лесной пищи жесткой и более сухой травянистой, вызвал весьма важные изменения в образе жизни и строении скелета непарнокопытных животных и определил следующий решающий этап в их эволюции к современным однокопытным формам. "Когда трехпалые анхитерий, - писал В. О. Ковалевский, - пошли на большие сухие луга древнего миоцена, то нужны были ноги только для опоры на сухой, твердой, невязкой почве; пошло развитие лошади и преобладание одного пальца и приспособление Gebiss'a (зубов) к травоядению*..."
* (В. О. Ковалевский, Палеонтология лошади, изд. Академии наук СССР, 1948, стр. 297.)
С переходом на травянистую пищу у предков лошадей, так же как и у большинства прочих копытных животных, усложняется зубной аппарат, увеличивается прочность перетирающих поверхностей (развитие складок эмали и цемента), зубы приобретают высокие коронки и способность к постоянному росту (по мере стирания они выдвигаются из челюсти). Этим компенсируется быстрое стирание зубной поверхности, вызываемое жесткой травянистой пищей, захватываемой нередко вместе с землей и песком.
В. О. Ковалевский показывает на ископаемых черепах, как в связи с усложнением зубного аппарата изменяется также строение челюстей, кости которых приобретали большую массивность и удлинялись, в результате чего глазница и черепная коробка в ходе эволюции отодвигались назад, а лицевая часть черепа удлинялась.
Жизнь на открытых равнинах развивала способность к быстрому бегу. Более плотная почва равнин служит достаточной опорой для среднего пальца, и боковые пальцы, необходимые прежде для передвижения по рыхлому, вязкому грунту лесов, становятся в новых условиях ненужной обузой.
В этот период у предков лошади завершается редукция боковых пальцев, и образуются однокопытные формы с длинными и стройными конечностями. Блоковидные суставы ног приобретают большую прочность в результате образования валиков, входящих в бороздки противостоящих костей, что укрепляет суставы и предохраняет их от вывихов*.
* (См. В. О. Ковалевский, Палеонтология лошади, стр. 210.)
Этот период эволюции представлен рядом переходных ископаемых форм.
Обнаруженные в миоцене Северной Америки остатки парагиппуса (Parahippus) и происходящего от него меригиппуса (Meryhippus), невидимому, являются переходной формой от древних лесных к более новым формам животных, жившим на открытых пространствах. Они характеризовались еще большей редукцией боковых пальцев, чем их предшественники. Боковые пальцы у них настолько укоротились, что во время быстрого бега, вероятно, уже не касались земли. Зубы отличались большей сложностью рисунка и большей длиной, однако не имели еще постоянного роста*.
* (См. А. Ш. Ромер, Палеонтология позвоночных, 1939.)
В верхнем миоцене и плиоцене Старого Света и Северной Америки получила также широкое распространение трехпалая форма лошади, описанная под названием гиппариона. Гиппариона долгое время считали (в том числе и В. О. Ковалевский) непосредственным предшественником лошади, однако позднейшие исследования и находки других, более близких к лошади форм, живших одновременно с гиппарионом, заставили признать гиппариона боковой ветвью, не оставившей последующих форм.
По развитию зубов, более сложному и своеобразному рисунку зубной эмали гиппарион как бы обогнал эволюцию основной линии лошадей своего времени, по строению же ног отстал от нее, сохраняя примитивные трехпалые конечности.
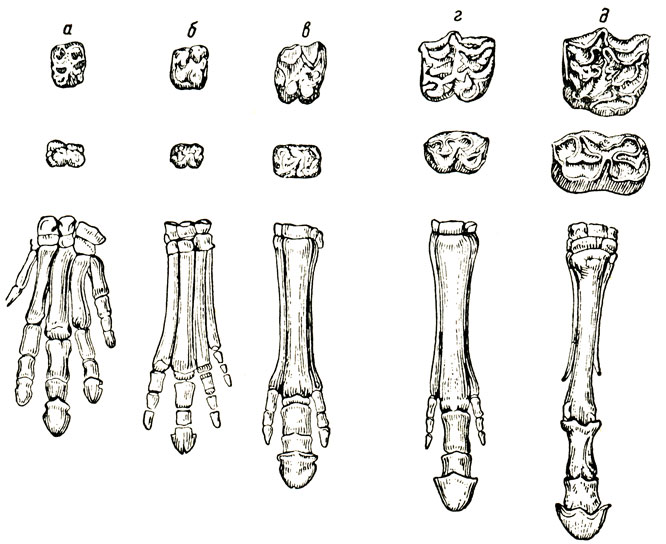
Рис. 84. Изменение зубов и ног у эквидов: а - один из видов фенакода (Tetraclaenodon); б - хиракотериум (Hyracotherium); в - анхитерий; г - плиогиппус; д - современная лошадь (по проф. М. В. Павловой)
Современные гиппариону лошади представляли собой уже вполне однопалую форму с рудиментарными косточками вместо боковых пальцев. К концу плиоцена гиппарион повсеместно вымер.
Следующие за описанными формы основной линии однокопытных уже обнаруживают в своем скелете признаки приспособленности к жизни на равнинах и употреблению жесткой пищи. Эти формы стоят весьма близко к современным лошадям. К их числу относится распространенный в плиоцене плиогиппус (Pliohippus). Это уже довольно крупное животное, с еще более редуцированными боковыми пальцами. От боковых пальцев у ряда форм остались только рудименты в виде грифелевидных косточек. Коренные зубы имели постоянный рост, были высококоронковыми, с более сложными, чем на зубах протогиппуса, складками*.
* (См. проф. М. В. Павлова, Палезоология, ч. II, 1929.)
В верхнем плиоцене и в эпоху, переходную от плиоцена к четвертичному периоду*, однопалые формы древних лошадей уже получили широкое распространение на огромных степных территориях Европы, Азии и связанных тогда еще с ними перешейками материков Северной Америки и Африки, часто встречаясь вместе с гиппарионом. Это был период возраставшего похолодания и уменьшения влажности, климата, характерный развитием в степной зоне специфической степной флоры и фауны. Именно в этот период происходит замена гиппариона-жителя саванн - однопалой лошадью - жителем сухих степей.
* (Четвертичный период состоит из эпохи плейстоцена (начавшейся около миллиона лет назад) и современной эпохи. Плейстоцен в свою очередь подразделяется отдельными учеными на нижний (самый древний), средний и верхний. Выделяют еще самую раннюю эпоху четвертичного периода (переходную от третичного) под названием постплиоцена.)
Древние лошади верхнего плиоцена, остатки которых обнаружены во многих местах Европы, Азии, Африки, Америки, в том числе и на Азовском побережье (Рютимейером, В. Громовым и др.), хотя и имели уже вполне однопалые конечности, но еще отличались от современных лошадей рядом существенных признаков, в частности более узколобой и длинномордой формой черепа, более складчатым строением и более мелкими размерами зубов и тонким костяком длинных конечностей.
Эта широко распространенная форма, европейские и африканские остатки которой описаны под названием "лошади Стенопа", явилась прародителем позднейших форм (в Америке представители аналогичной формы описаны под названием Е. Plesippus).
Плиоценовая лошадь Стенона обнаружена в виде двух разновидностей: более мелкой, типичной формы - Е. Stenonis tipicus и более крупной, приближающейся по росту к самым крупным тяжеловозам, но легкой по сложению и тонконогой, которая описана под названием Е. Stenonis major*.
* (См. В. Громова, Опыт изучения процесса образования форм у млекопитающих (род Equus лошади). Труды зоологического института Академии наук СССР, т. VI, вып. 4, 1941, стр. 124.)
Близкой по сложению к Е. Stenonis являлась плиоценовая лошадь Китая Е. Sanmeniensis, а также найденная в Индии форма Е. sivalensis.
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://konevodstvo.su/ 'Konevodstvo.su: Коневодство и коннозаводство'