Глава II. Происхождение и одомашнивание лошадей
Происхождение лошадей. По зоологической классификации домашняя лошадь, ее дикие предки, вымершие и ныне живущие сородичи относятся к отряду непарнокопытных (непарнопалых), к семейству лошадиных (Equidae), к роду лошадей (Equus). Род лошадей объединяет четыре подрода: настоящих лошадей (Е. Equus), полуослов (Е. Hemionus), ослов (Е. Asinus) и зебр (Е. Hippotigris), каждый из которых включает по нескольку видов.
Представители разных родов не скрещиваются, а животные одного рода, но разных перечисленных выше подродов при скрещивании между собой дают бесплодное потомство. Исключение составляют лишь гибридные самки, случаи плодовитости которых описаны в литературе. Бесплодие гибридных самцов объясняется нарушением сперматогенеза, что обусловлено различиями в кариотипе скрещиваемых животных разных подродов. Обнаружены различия в количестве хромосом: у домашней лошади (2п) их 64, лошади Пржевальского - 66, кулана - 54, онагра - 56, зебры Гранта - 52 и у осла - 62. Следует отметить, что причины бесплодия гибридных животных, полученных в результате скрещивания представителей разных подродов, еще недостаточно выяснены.
Изучение происхождения лошадей до недавнего времени основывалось преимущественно на находках древнего изобразительного искусства. Затем исследователи сосредоточили свое внимание на изучении костных остатков животных, обнаруженных во время палеонтологических и археологических раскопок. В настоящее время, применяя метод радиокарбонного анализа, удалось более точно установить возраст этих находок, который оказался значительно большим, чем предполагалось ранее.
В соответствии с современными представлениями эволюция рода лошадей проходила на протяжении около 70 млн. лет. Понять длительный процесс исторического развития органических форм стало возможно благодаря эволюционному учению Ч. Дарвина, изложенному им в работе "Происхождение видов" (1859 г.) и принесшему автору мировую славу. Ч. Дарвин доказал, что движущими силами эволюции животных и растений являются изменчивость, наследственность и естественный отбор, результате наследственной изменчивости и естественного отбора на протяжении многих поколений возникали новые группы животных, отличавшиеся от своих предков. Классическим примером этого служит история происхождения лошадей, в разработку которой неоценимый вклад внес выдающийся русский ученый В. О. Ковалевский (1842 - 1883).
Благодаря В. О. Ковалевскому эволюционная теория Ч. Дарвина стала той теоретической основой, на которой быстро развивались палеонтология и палеоботаника, а также возник ряд новых направлений, в частности учение об адаптациях в прошедшие геологические периоды - палеоэкология.
В. О. Ковалевский - первый палеонтолог, широко применивший эволюционное учение Ч. Дарвина для решения конкретных вопросов эволюции позвоночных на материале копытных. Он установил наличие адаптивных и инадаптивных путей эволюции в палеонтологической истории непарнокопытных. Углубленный анализ преобразований костей конечностей животных показал, что в разных филогенетических рядах эти преобразования носили неодинаковый характер. В одних случаях кости запястья и предплюсны при прогрессивном развитии средних пальцев и редукции крайних сохранили свое расположение в ряд. В других случаях произошла их перегруппировка, что придало определенную прочность конечностям и обеспечило приспособленность животных к быстрому бегу. Первый путь оказался инадаптивным. Шедщие по этому пути эволюции группы животных чаще всего нe выдерживали конкуренции в борьбе за существование и вымирали. Во втором случае они оказались лучшe приспособленными и дали в дальнейшем современных непарнокопытных, в том числе лошадиных. Эти исследования имели большое значение для понимания причин вымирания многих филогенетических ветвей. Вместе с тем они явились блестящей иллюстрацией относительности органической целесообразности в эволюции.
Изученная эволюция ископаемых предков лошади имеет еще существенные пробелы и неточности, но она убедительно свидетельствует о превращении лесных стопоходящих животных ростом 30 - 45 см в крупных однопалых обитателей степей (рост 90 - 120 см - меригиппус). За прошедшие миллионы лет в связи со значительными изменениями геолого-климатических и растительных факторов ареала ископаемых предков лошадей естественный отбор устранил многие формы, оказавшиеся в конкретных условиях маложизнеспособными, и сохранил наиболее приспособленные. На протяжении почти всего третичного периода лошадь прошла исключительно сложный путь эволюции, выразившийся в крупных последовательных изменениях в строении всего организма, в особенности конечностей и зубов, обеспечивающих лучшую приспосабливаемость животных к условиям среды обитания.
В процессе эволюции отмечалось непрерывное появление малых и крупных мутационных изменений в фенотипе лошади, а естественный отбор способствовал накоплению полезных для вида мутаций и уничтожению особей - носителей мутаций, вредных для жизнеспособности популяции. Внутривидовая дифференциация обусловливалась в первую очередь изменениями фенотипа представителей вида под влиянием мутационных процессов, то есть генетически. Основную роль в эволюции лошадей сыграли множественные малые мутации, не вносящие заметных изменений в фенотип животных, но накапливающиеся в популяции в гетерозиготном состоянии. Крупные мутационные изменения у лошадиных в гомозиготном состоянии чаще всего вели к появлению маложизнеспособных форм, устранявшихся отбором. Здесь уместно привести слова И. И. Шмальгаузена, который писал, что эволюция обусловливается многообразием генетической структуры вида и разнообразием условий его существования. Схема эволюции лошади приведена на рисунке I.
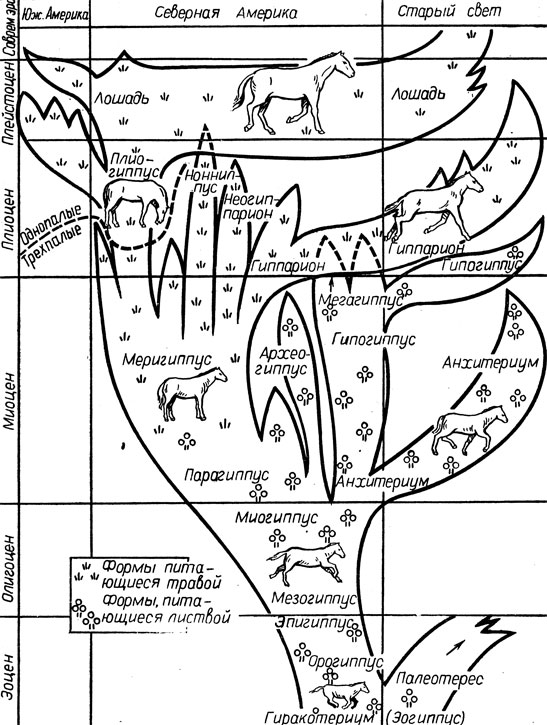
Рис. 1. Схема эволюции лошади (по данным В. О. Витта, О. А. Желиговского и др., 1964)
Ныне ученые начинают палеонтологическую историю лошади с рода Хиракотериум (Hyracoterium). В Западной Европе Хиракотериум, а в Северной Америке Эогиппус были животными небольшого роста (высота в холке от 25 до 56 см), с четырьмя хорошо развитыми пальцами на передних ногах и с тремя пальцами на задних ногах. Зубы Эогиппуса имели невысокую коронку, а коренные зубы были приспособлены для пережевывания сочной и мягкой пищи (листья, плоды).
Появление мезохиппуса в нижнем олигоцене характеризует следующий этап в эволюции рода Эквус (лошадь). Эти животные имели на всех ногах по три пальца, опирались на них при ходьбе, на зубах у них возникли эмалевые гребни. Рост животных был 45 см. Таким образом, более 30 млн. лет, прошедших со времени эпохи эоцена до олигоцена, мало внесли изменений по сравнению с предшествующими формами.
В эпоху миоцена некоторые ветви рода лошади оказались в резко изменившихся условиях жизни (в степи) и были вынуждены питаться степной, сухой и жесткой растительностью, что ускорило процесс адаптации животных. В миоцене появляется мерихиппус ростом 90 - 120 см, опиравшийся главным образом на один средний палец из трех, снабженный копытом. Зубы сильно изменились: удлинились коронки, появились мощные складки эмали и прослойки цемента, что обеспечивало пережевывание более сухой и грубой пищи. Многие боковые ветви (например, анхитериум), хуже приспособленные к новым условиям, вымирали.
В эпоху плиоцена появляются первые однопалые предки лошади - плиохиппусы, потомки которых были широко распространены по всем частям Света, вытеснив трехпалых эквид, происходивших от мерихиппуса, в том числе и гиппариона. Плиохиппус - крупное животное с редуцированными первым, вторым, четвертым и пятым пальцами, типичный обитатель сухих степей. Коренные зубы имели высокую коронку, значительные отложения цемента в углублениях между развитыми складками и гребнями эмали.
Трехпалый гиппарион появился в эпоху верхнего миоцена и широко распространился в плиоцене на таких континентах, как Америка и Евразия, но, не выдержав конкуренции с однопалым предком лошадей - плиохиппусом, вымер, несмотря на то что по развитию своих коренных зубов стоял на более высокой ступени эволюции, чём плиохиппус. Гиппарион представляет собой вымершую боковую ветвь генетического ряда рода Эквус, а поэтому не может считаться одним из предков современной лошади. Р. Стертой (1942), Г. Симпсон (1945), В. Громова (1949) и другие убедительно опровергают мнение тех палеонтологов, которые считают гиппариона прямым предком современной лошади.
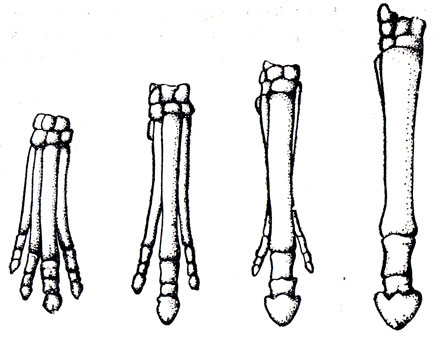
Рис. 2. Развитие передних конечностей в историческом ряду лошадиных. Слева направо: хиракотериум, мезохиппус, мерихиппус, лошадь (по данным Ренча, 1956)
Однопалые предки диких лошадей впервые появились в Северной Америке (рис. 2). В верхнем плиоцене многих мест Европы, Азии, Африки и Америки обнаружены останки древних лошадей, хотя и имевших однопалые конечности, но существенно отличавшихся от современных лошадей более узколобым и длинномордым черепом, складчатым строением мелких зубов и более длинными конечностями с тонкими костями. Представителем этой формы является Е. Stenonis, обитавшая в Европе и Африке, а в Америке - Е. Plesihippus. Близки по телесложению к Е. Stenonis древние лошади, останки которых найдены в плиоцене Китая (Е. Sanmeniensis) и Индии (Е. Sivatensis).
В течение четвертичного периода Европа перенесла четыре оледенения, вызвавших сильные изменения в растительном и животном мире. На основании изучений скелетов ископаемых лошадей считают, что род Эквус возник, по-видимому, в Америке и характеризуется представителями подрода Plesihippus, а древнейшие представители рода лошади в Старом Свете (Е. Stenonis) очень близки, а может быть, даже тождественны американскому подроду плезихиппус.
В Америке было найдено больше ископаемых останков предков лошадей, а так как в третичном периоде Северная Америка была соединена перешейком с Азией (на месте Берингова пролива), то предполагают, что лошади произошли в Америке и потом переселились в Азию, Европу и Африку. Вследствие сплошного оледенения Северной Америки дикие эквиды там погибли и до XV в. ни диких лошадей, ни других эквид в Америке не было.
В верхнем плиоцене (5 млн. лег назад) Старый Свет был заселен первыми однопалыми лошадьми, которых в это время не было в Австралии, может быть, их не было даже в южной Азии и на крайнем севере Евразии. Эти предки лошади вытеснили предшествовавших им в большом количестве гиппарионов, частично сосуществуя сними некоторое время.
Такая замена биологически господствующего семейства была связана, очевидно, с изменением климата и растительности на большей части ареала.
Переход от саванного ландшафта с более сочной растительностью, со сравнительно обильным увлажнением почвы к сухим степям благоприятствовал победе однопалых лошадей с их гипсодонтными (увеличенной высотой коронок коренных зубов по сравнению с их горизонтальными размерами) зубами и толстой эмалью над трехпалыми гиппарионами с более низкой зубной коронкой и более тонкой эмалью (В. Громова, 1949).
Определяющими развитие рода лошадей являются, очевидно, различие физико-географических условий, быстрота и степень их изменений. В Европе в плейстоцене, где имело место наиболее резкое изменение климата и ландшафта, сложился подрод настоящих лошадей, сильнее всего отклонившийся от исходных форм плиоценовых предков.
В Африке, Центральной и южной Азии с их более постоянными условиями климата и растительности сохранились архаические формы - подроды зебр, ослов и полуослов. Но и в Европе в специальных, особо ксерофит-ных условиях удержались более архаичные формы.
С раннего плейстоцена подрод истинной лошади можно считать сложившимся и все представители его здесь принадлежат к одному виду настоящей, теперь домашней лошади, но к разным его вариететам. Степной ландшафт в одних случаях выступал в роли фактора, ускоряющего эволюцию, а в других случаях - задерживающего эволюцию, сохраняющего архаичность их форм (например, полуослы и лошадь Пржевальского).
При переходе от плиоцена к плейстоцену такие факторы, как увлажнение и облесение, вызвали быстрое превращение Е. Stenonis в Е. Caballus (подрод Allohippus в подрод Equus).
В позднем плейстоцене Европы значительное изменение форм лошадей происходило в связи с эпохами похолодания, когда появились более крупные и массивные ширококостные (широконогие) формы, а в эпохи потеплений- более легкие и тонконогие формы.
О генетической связи разных вариететов между собой можно предполагать, но конкретные представления составить мы не можем, так как имеющиеся данные разрознены.
Ныне всех лошадей Европы и Северной Африки, начиная с раннеплейстоценовых, обычно объединяют в один вид Е. Caballus L., включая диких и домашних. Поступают так потому, что видовой состав диких лошадей Европы к периоду их одомашнивания неизвестен, а также потому, что остеологические различия на грани диких и домашних лошадей установить очень трудно.
В Северной Америке эволюция рода Equus достигла стадии Е. Plesihippus, а настоящие лошади в диком состоянии не обнаружены. В Северной Америке лошади появились только в XV в. н. э. после завоза их из Европы. Отмеченная В. Громовой общая тенденция к укрупнению плейстоценовых ископаемых лошадей Европы в периоды похолоданий и уменьшению их формата в периоды потеплений недостаточна для установления генетической связи между современными породами лошадей и их древними предками эпохи плейстоцена.
В Европе, Азии и Африке под влиянием различных условий жизни возникли новые формы эквид, из которых в дальнейшем сформировались существующие до наших дней четыре подрода Equus: ослов, полуослов, настоящих лошадей и зебр.
В диком состоянии сохранилась лишь лошадь Пржевальского (рис. 3), а другой представитель дикой лошади- тарпан был истреблен еще в прошлом веке. Полуослы и зебры существуют только в диком, а ослы - в диком и одомашненном состоянии.
Тарпаны (Е. Caballus Gmelini Antonius) водились в южнорусских степях до 70-х годов XIX в. С. Гмелин писал: "...лет двадцать тому назад (то есть в 1748 г.) здесь по соседству с Воронежом много было диких лошадей." В 1768 г. С. Гмелин добыл дикого жеребца тарпана и описал его. Некоторые ученые (Антониус, Громова и др.) полагают, что тарпан представляет собой самостоятельный европейский вид дикой лошади. К сожалению, палеонтологи располагают всего одним скелетом и двумя черепами южнорусского тарпана. Череп тарпана принципиально ничем не отличается от черепа наших лошадей, однако величина его и высота затылка меньше. Но тарпан не был потомком обитавших до него на восточноевропейской равнине позднеплейстоценовых лошадей, которые, по-видимому, переселились к северу в лесную зону, уступив свое место уже в недавнее время иммигрантам (тарпанам) из юго-западной Азии не ранее неолита южной полосы Восточной Европы. Очень вероятно, что кровь тарпана течет во многих лошадях русских пород.

Рис. 3. Лошади Пржевальского в степи Аскания-Нова
Рост тарпанов достигал 135 см. Голова у него была большой, с прямым профилем; шея -короткой и толстой; конечности - сухими. На передних конечностях слабо развитые каштаны. Масть тарпанов мышастая, с темным "ремнем" по спине.
Современные представители рода Equus. К семейству лошадиных относят собственно лошадей, полосатых лошадей (зебр), полуослов и ослов. Одни авторы полагают, что в этом семействе следует выделить три рода (ослов, зебр и лошадей), а другие, в том числе В. О. Витт, В. Громова и С. Н. Боголюбский, включают в семейство лошадиных только один род (Equus), подразделяя его на четыре подрода: зебр, ослов, полуослов и лошадей (эквус кабаллус), убедительно аргументируя свою точку зрения возможностью плодовитого скрещивания всех представителей данного семейства. Каждый из подродов характеризуется немногими отличительными признаками, развившимися у них уже после отделения от главного ствола Equus и от ветвей этого ствола. Существуют также внешние и экологические отличия всех четырех подродов, однако этого мало для выделения каждого из них в самостоятельные роды.

Рис. 4. Зебра Чапмана с 10-месячным гибридным жеребенком Рыжиком (слева), родившимся в 1973 г. в Ленинградском зоопарке (мать - зебра, отец - кулан)
Подрод зебр (полосатые лошади Африки) представляет наиболее архаическую и менее всего изменившуюся от плиоценового предка ветвь. Известно несколько видов зебр, различающихся не только по строению скелета, черепа и росту, но и по расположению черных полос на теле (зеброидностью). Живут зебры табунами, они плохо акклиматизируются и трудно приручаются. Среди разновидностей зебр наиболее известны небольшие горные, а также крупные саванные зебры Греви. В "Аскании-Нова" имеются зебры (рис. 4), в том числе Греви и Чапмана. С лошадьми зебры дают бесплодное потомство. В 1973 г. в Ленинградском зоопарке родился гибрид Рыжик. Он был получен в результате скрещивания зебры Чапмана с самцом куланом. Продолжительность беременности зебры составила 368 дней.
Подрод ослов, так же как и зебры, представляет самостоятельную ветвь рода лошадей, отделившуюся от других ветвей уже с плиоцена и несколько отставшую от них в своем дальнейшем развитии. Дикие ослы встречаются только в Африке в двух близких разновидностях: сомалийской и абиссинонубийской, которые формировались в сравнительно малоизменявшихся климатических условиях. Это отразилось на биологических особенностях животных, в частности на периоде плодоношения, который у ослиц составляет 12 месяцев (365 дней), выжеребка происходит не в строго определенное время года.
Голова ослов большая, с длинными ушами, челка отсутствует, грива короткая; туловище длинное, с низкой холкой, слабо омускуленным крышеобразным крупом и коротким хвостом, оброслость которого удлиненными волосами начинается с нижней его трети. У ослов узкие и высокие копыта. Ослы издают характерные звуки. В странах Востока ослов используют издавна в качестве рабочих и транспортных животных, причем раньше, чем лошадей. Существует большое число пород, сильно варьирующих по росту (80 - 150 см), но все они происходят от диких ослов Африки. При скрещивании с представителями рода лошадей ослы дают потомство, в котором самцы всегда бесплодны. Животное, полученное в результате скрещивания кобылы с ослом, называется мулом, а от ослицы и жеребца - лошаком. Мулы крупнее лошаков, отличаются высокой работоспособностью и долговечностью. Мулопроизводство имеет и в наши дни определенное народнохозяйственное значение в ряде стран, где получают крупных упряжных, вьючных и верховых мулов от кобыл и ослов соответствующего типа путем целенаправленного подбора их для скрещивания.
Подрод полуослов представлен несколькими видами, из которых кианг - самое крупное животное (высота в холке около 130 - 140 см); кианги обитают на плоскогорьях Юго-Западного Китая и Тибета. Онагр встречается редко, причем только в диком состоянии в Иране, Афганистане и юго-восточной части Туркменской ССР. Высота в холке онагра 116 - 130 см. У киангов по сравнению с онаграми масть темнее. Рост кулана 125 - 137 см. Он больше похож на лошадь, чем на осла.

Рис. 5. Кулан самка с куланенком в 'Аскании-Нова'
Отличительные особенности экстерьера куланов: легкость и стройность, длинные и сухие конечности, более короткие, чем у ослов, уши, хвост ослиного типа, копыта по форме занимают промежуточное положение между лошадиными и ослиными (рис. 5). Каштаны имеются только на передних конечностях, "щетки" слабо развиты. Куланы, как и другие представители подрода полуослов, относятся к быстро исчезающим диким формам из семейства лошадиных, встречающихся в сухих степях, полупустынях и на плоскогорьях Азии (Монголия, Китай, Индия, Афганистан). В СССР организованы заповедники куланов (онагров). Весьма важной особенностью куланов является их способность к быстрому бегу (до 85 км/ч) и выносливость.
Некоторые исследователи считали, что подрод полуослы имеет с лошадью больше общего, чем ослы, и на этом основании полагали возможным получить плодовитых гибридов от скрещивания кулана с лошадью. В 1933 г. по инициативе В. О. Витта и В. А. Щекиаа Ташкентская зональная опытная станция начала работу в этом направлении, но только в мае 1936 г. удалось выловить 5-дневного куланенка-самца, которого вырастили в одном стойле с ослицей того же возраста. Полностью приручить этого кулана не удалось. Летом 1938 г. кулан был перевезен в Ташкентский конный завод, где его спермой осеменили кобыл и ослиц, давших несколько десятков гибридов - конекуланов и ослокуланов, оказавшихся бесплодными. Продолжительность плодоношения от кулана у кобыл в среднем составила 339 дней, а у ослиц - 346.
Подрод лошадь (Equus Caballus) включает быстро исчезающий вид диких лошадей Пржевальского и весьма разнообразную и многочисленную группу домашних лошадей.
Дикая лошадь Пржевальского была широко распространена в степной и безводной зоне Центральной Азии, а в наше время сохранилась в небольшом количестве в Монголии и на Украине в заповеднике "Аскания-Нова". Рост этой лошади небольшой (124 - 135 см). Голова грубая, с короткими ушами, без челки, но с длинными волосами (баками) под нижней челюстью; шея массивная, короткая; холка низкая; конечности тонкие, копыта широкие, имеются каштаны. Масть саврасая различных оттенков, вдоль спины проходит темный узкий ремень, низ конечностей черный. Грива стоячая, черная, как и хвост, покрытый у корня короткими волосами, удлиняющимися книзу. Лошадь Пржевальского ржет и храпит, как и домашняя лошадь, жеребость продолжается 340 - 350 дней с выжеребкой в апреле - мае, при скрещивании с домашней лошадью дает плодовитое потомство.
Мнение о том, что лошадь Пржевальского явилась непосредственным диким предком домашней лошади, не подтвердилось исследованиями В. Громовой, Б. Ф. Румянцева, Б. П. Войтяцкого и других, которые считают ее самостоятельной ветвью лошадей Центральной Азии. Диких лошадей, кроме лошади Пржевальского, не сохранилось. Мустанги США представляют собой одичавших лошадей, а в Австралии одичавшие лошади известны под названием брамби.
Одомашнивание и использование лошадей. Одомашнивание (доместикация) лошадей началась в глубокой древности. Первобытному человеку потребовалось длительное время для постепенного познания особенностей жизни и поведения диких лошадей, а затем освоения приемов охоты на них для удовлетворения своих потребностей в пище, а в дальнейшем и приручения. С. Н. Боголюбский и другие полагают, что приручение диких лошадей было процессом очень длительным, оно проходило на протяжении не менее 500 тыс. лет. Считают, что различие между приручением диких лошадей и последующим их одомашниванием заключается в том, что приручение всегда начиналось с отдельных особей и часто прекращалось на них, а доместикация характеризовалась массовостью и непрерывностью процесса, требующего смены нескольких поколений животных в популяциях. При этом отличия домашних лошадей от диких возрастали.
В известной мере сохраняет значение мнение многих ученых о том, что основное отличие прирученных лошадей от одомашненных заключается в том, что прирученные животные не размножаются в неволе, а одомашненные размножаются. Однако в последнее время человек научился получать потомство от многих диких животных, находящихся в неволе (например, в зоопарках), в том числе и от таких диких родичей домашней лошади, как от лошади Пржевальского и кулана.
В настоящее время установлено, что процессы приручения и одомашнивания представителей семейства лошадиных происходили независимо в различных первичных очагах много раньше, чем предполагали еще недавно. Возраст древних поселений людей Передней Азии достигает VIII, а может быть, и IX тысячелетия до н.э. Земледелие и животноводство на Древнем Востоке, по-видимому, возникли до начала существования Египта.
Н. И. Вавилов называл два основных очага одомашнивания лошадей: юго-западноазиатский и средиземноморский. Приручение и одомашнивание диких лошадей первоначально осуществлялись в нескольких степных регионах Восточной Европы и в Азии, что привело к появлению разновидностей домашних лошадей на территории их распространения. В. О. Витт считал бесспорным, что крупные формы предков лошади к концу плейстоцена уступили место пришедшим со стороны менее крупным, опровергая мнение Герре, считавшего причиной измельчания крупных видов их вымирание. Учитывая длительность и сложность процесса доместикации лошадей, надо отметить, что очагов их первичного одомашнивания было немного, а дальнейшее распространение происходило с использованием накопленного людьми опыта приручения животных и заимствованием уже одомашненных лошадей различными племенами и народами. .
Одним из первых очагов доместикации лошадей обычно называют обширную территорию Средней Азии, прилегающую к pp. Амударье и Сырдарье. Очевидно, здесь были приручены и одомашнены дикие лошади, имеющие стройное телосложение. Одомашнивание лошадей происходило также в лесных районах Азии и Европы. В Якутии и на Урале найдены останки диких и домашних лошадей, отличавшихся от степных. По-видимому, эти регионы относятся ко вторичным очагам доместикации лошадей.
В. О. Витт предполагал, что предки домашней лошади существовали в районах с умеренным климатом, со степным ландшафтом, к которому они биологически больше приспособлены, чем дикие ослы. По мнению В. Громовой, В. О. Витта и других исследователей, лошади в течение плейстоцена и голоцена были распространены в субарктических, а ослы и зебры - в субтропических районах. Сказанное в известной мере объясняет и то, что ослы и онагры были одомашнены раньше лошадей, доместикацию которых относят к концу неолита - нового каменного века, то есть позднее одомашнивания собак, овец, коз и крупного рогатого скота. Знаток истории домашних животных В. И. Цалкин отмечал, что современные представления о месте и времени одомашнивания лошадей лучше документированы, чем для других домашних животных. В то же время в этом направлении необходимы дальнейшие исследования.
Выявлена определенная последовательность в использовании лошадей. Началось оно с охоты первобытного человека на диких лошадей с целью обеспечения себя мясом, молоком, а позднее и кумысом. Включение в пищу кобыльего молока стало возможным после длительной работы человека по приручению лошадей и характерно для перехода наших предков от собирательства и охоты к пастушеству, а также для племен и народов, стремившихся к оседлости и земледелию, которые длительное время сочетались с кочевым и отгонным животноводством. Для перевозки несложного имущества людей сначала использовали крупный рогатый скот, а затем ослов и лошадей.
Сложнее обстоит дело с определением последовательности и времени использования лошадей в упряжи, в повозке, под седлом и вьюком. К сожалению, археологические, этнографические и культурно-исторические памятники, свидетельствующие о значении лошади в хозяйстве человека и в социально-историческом развитии общества, относятся к довольно позднему времени. Этим объясняется и утверждение того, что доение кобыл применялось скифами только в V в. до н. э., хотя маловероятно такое позднее включение кобыльего молока в пищу человека. Есть данные о том, что кочевники в I тысячелетии до н. э. умели приготовлять кумыс и высоко ценили его достоинства.
Одомашнивание лошадей позволило человеку использовать их в различных целях в зависимости от социальных и природных условий, а также от направления своего хозяйства. Очень хорошо было развито коневодство у кочевых народов, для которых с древнейших времен на протяжении тысячелетий оно являлось основой хозяйства, а достигшие высокого уровня развития культуры такие государства, как Ассирия, Вавилон и Египет, примерно до начала II тысячелетия до н. э. коневодством не занимались. Лошадь появилась в этих государствах лишь за 2 - 1,5 тыс. лет до н.э., после их завоевания кочевыми народами. В. Б. Ковалевская упоминает о древней китайской летописи, в которой говорится о легкой коннице скифов и саков, гуннов и авар, тюрок и монголов, "бушующих, как бури и молнии", не знающих устойчивых боевых порядков, наводящих страх и панику на врага.
Рано поняв большое значение лошади в хозяйстве, люди стали уделять все больше внимания коневодству, переросшему к концу IV тысячелетия до н. э. в своеобразный культ коня у различных племен, в том числе и населявших нашу территорию. Кочевники использовали лошадей под верхом, сначала без седла и уздечки, о чем свидетельствуют изображения всадников на монетах III в. до н. э. и наскальные рисунки. Если нумидийские всадники и ливийская конница обходилась без уздечки и удил, то возможность такого использования лошадей в коннице кочевых племен и народов была значительно раньше.
Из обширных евразийских степей кочевые народы проникли в страны Древнего Востока и Китай, куда принесли с собой примерно за 2 тыс. лет до н. э. опыт разведения лошадей и использования конницы в военном деле. Существует и другое мнение, что кочевники в странах Древнего Востока познакомились с использованием крупного рогатого скота в упряжи, переняли этот опыт, перенеся его в коневодство. Использование лошадей в боевых колесницах было широко распространено в государствах Малой Азии, а затем в Древней Греции и Риме, что усиливало культ коня. О культе коня в V в. до н. э. писал Геродот, но в рабовладельческих государствах лошадь как тягловая сила на сельскохозяйственных работах еще не применялась.
Уже за 1 тыс. лет до н. э. в некоторых государствах (Хорезм, Бактрия и др.) были выведены легкие, сухие, быстроаллюрные лошади. Древняя Бактрия, по словам В. О. Витта, была хорошо известна окружающим народам процветающим коневодством. Первыми очагами образования древних специализированных пород верховых лошадей считают Мидию и Персию, впоследствии давших туркменских и персидских лошадей.
В Центральной и Северо-Западной Европе долгое время использование лошадей не было специализированным. Была распространена преимущественно небольшая лесная лошадь, которую на сельскохозяйственных работах стали систематически применять в XI - XII столетиях. Только при феодальном строе, в период средневековья, начался процесс специализации типа лошадей в направлении их укрупнения под всадника - тяжеловооруженного рыцаря, весившего около 200 кг. Опыт боевых столкновений рыцарей на тяжелых лошадях с легкой и подвижной конницей восточных народов, особенно после изобретения пороха, резко изменил требования к кавалерии. Она нуждалась в легких и резвых лошадях, которые и были получены в результате скрещивания местных лошадей с лучшими восточными. Так, в Европе появились новые породы верховых лошадей, классическим представителем которых стала чистокровная верховая порода, выведенная в Англии путем сложного воспроизводительного скрещивания при использовании восточных лошадей.
Значительные преобразования произошли не только в промышленности и земледелии, но и в животноводстве Западной Европы с развитием капитализма, начало которого относится к XVI в. Страной, раньше других ставшей на путь капиталистического развития, стала Англия, где происходила интенсивная дифференцировка и специализация типа и пород сельскохозяйственных животных, в том числе и лошадей. Для колониальных войн совершенствовался тип верховых лошадей, а для работ на транспорте и в сельском хозяйстве потребовались крупные упряжные кони, основой выведения которых послужила уже созданная ранее рыцарская лошадь. В этот период в Англии было выведено несколько пород (шайры, клейдесдали, суффольки) - животные, которые отличались большой живой массой. Появились бельгийские тяжеловозы, а несколько позже в России - крупные упряжные лошади.
Рысистые породы лошадей были созданы не только в Европе (голландская, орловский рысак), но и в Северной Америке (американский рысак). Итак, процесс дифференциации и специализации пород лошадей, расширение их ареала проходили в конкретных социально-экономических и естественноисторических условиях различных зон земли.
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://konevodstvo.su/ 'Konevodstvo.su: Коневодство и коннозаводство'