Век иной - иные кони...

Сегодня, восхищенно провожая глазами несущийся в грохоте копыт и волнах пыли табун лошадей, создающий переливающимися бликами мышц и развевающимися хвостами ощущение полета, мы невольно спрашиваем себя: что это за чудо? Откуда появился этот властелин времени и пространства? Что за прародитель создал такое волшебное создание - лошадь, в каждой черточке и движении которой видится свист степных ветров, радость жизни, гармония и очарование...
Для того чтобы узнать, почему лошадь так идеально приспособлена к жизни на открытых пространствах и находится в гармоничном единстве с равниной, нам нужно проследить историю рода лошадей во времени, на протяжении миллионов лет. Но как заглянуть в прошлое, не имея машины времени? На помощь нам приходит эволюционная палеонтология.
Происхождение жизни и возникновение более сложных живых форм из более простых явилось ответом сначала химических соединений, а затем органических образований на изменяющиеся внешние условия. Когда-то возникшие благоприятные условия среды дали жизнь первым зачаткам живого на планете, которые, появившись на свет, уже "не хотели" вернуться в никуда. Использовав эти условия для накопления и закрепления приспособительных качеств, эти первые организмы начали активно размножаться, дав начало гонке, которая не останавливается, а лишь убыстряется и по сей день. В чем ее смысл?
Прежде всего смысл в том, что любое живое стремится выжить, выжить во что бы то ни стало и дать потомство. Условия среды, в которых оно находится, никогда не бывают постоянно благоприятными, а, наоборот, очень изменчивы, поэтому все живое стремится воспроизвести как можно более многочисленное потомство, вооруженное к тому же приспособлениями (адаптациями) к жизни в конкретной меняющейся среде. Поскольку же формы, в которых выступает физическая материя на планете, очень многообразны, великое разнообразие растений и животных отражает адаптивную реакцию организмов на колебание условий среды на протяжении тысячелетий.
Внутри каждого конкретного вида живых существ, будь то насекомое, растение или млекопитающее, рождается множество в общем похожих организмов, отличающихся небольшими различиями в строении тела, функциях разных органов и поведении, т. е. генетическая изменчивость вида поставляет материал для эволюции. Факторами эволюции на этой ступени служат изменения в генах (мутации), их частота и характер, продолжительность жизни поколений (чем быстрее смена поколений, тем выше скорость эволюции), размер популяции (чем больше популяция, тем шире зона ее распространения, тем более изменчивы экологические условия, в которых она существует, тем выше изменчивость и скорость эволюции). Затем в "игру" вступает следующий эволюционный фактор - пресс среды. Под действием разнообразных условий конкретных биотопов происходит отсечение части или частей генетического потока. При этом выживает и оставляет себя в потомстве лишь наиболее приспособленный, лучше других усвоивший уроки жизни. И, наконец, незначительные на первый взгляд изменения, подкрепляемые направленным отбором, через время и через тысячи поколений живых существ приводят к появлению новых форм. Поэтому организмы, существующие в каждой конкретной эпохе и конкретном месте в пространстве, отражают совокупностью строения своих форм приспособляемость к имеющимся в данный момент условиям. При этом наметанный глаз исследователя зачастую видит в строении конкретного организма и его прошлое - через призму существующих адаптации к той геологической эпохе, которая предшествовала эпохе жизни этого организма. И наоборот, имея в руках конкретные формы живых существ (например, ископаемые останки), населявшие определенную часть земного шара в определенный период времени, можно по аналогии с тем, что известно о потребностях и функциях современных аналогов этих форм, делать выводы о климатических и геологических особенностях тех мест в давний период. Таким же образом можно заглянуть и в историю рода лошадей.
Архаичные лошадиные
Пытаясь найти предка лошади, можно сколь угодно долго путешествовать по эволюционной пирамиде, добравшись в конце концов до первых млекопитающих или даже дальше, до амебы. Но мы остановимся на животном, уже несшем в себе основные задатки, которые под влиянием определенных геологических катаклизмов и экологических условий развились на протяжении более чем 50 миллионов лет в то гармоничное и волнующее нас существо, которое мы называем лошадью.
Итак, прежде чем увидеть это животное, перенесемся в ранний эоцен - на 55 миллионов лет назад, на континент, который можно назвать Евро-америкой. В то время современная Европа и Северная Америка еще не были разъединены Атлантическим океаном, а от Азии их отделял морской пролив, пролегавший там, где сейчас находится Уральский хребет. Это был цветущий континент, с тропическим климатом, покрытый вечнозелеными лесами. Воздух был полон густых дурманящих испарений, а земля взращивала богатый урожай кустарников, лиан и тропических великанов-деревьев, уходящих стволами далеко ввысь. Там, где через густой полог кроны тропического леса пробивались солнечные зайчики, на лесных полянах жили необычные, ростом с собаку, млекопитающие, питавшиеся листвой кустарников и мягкими лесными травами,- гиракотериумы, или эогиппусы. У них были маленькая голова на короткой шее, длинное туловище и короткие лапы. На передних ногах эогиппусы имели по четыре, а на задних - по три пальца.
Низкие коренные зубы этих животных выглядели как четырехугольные костяные столбики с четырьмя-шестью бугорками на жевательной поверхности. Предкоренные зубы тоже были примитивны, с треугольной поверхностью. Конечно же, такими зубами можно было перерабатывать не жесткую степную растительность, а только сочные мягкие листья да мелких беспозвоночных. Слабость жевательной системы подтверждалась и коротким лицевым отделом. Глазницы находились посредине черепа.
Ноги у этого существа были полусогнутые и низкие. При ходьбе эогиппус опирался на подушечки пальцев, каждый из которых заканчивался коготком (будущие копыта). При передвижении животное напоминало современную собаку. Ясно, что с такими ногами нельзя было состязаться в скорости с быстроногими хищниками на открытых пространствах, поэтому эогиппус прятался в лесной чаще, зарослях кустарников и был "одет" в шкуру с коротким и редким волосяным покровом, полосатой или пятнистой окраски, подобно современным лесным и саванновым животным.
В среднем и верхнем эоцене Северной Америки встречаются останки форм, очень близких к описанной нами и отличающихся от нее сравнительно небольшими, но уже более прогрессивными изменениями. Так, предкоренные зубы уже приняли форму коренных, следовательно, истирающая поверхность щечных зубов несколько увеличилась.
Эогиппус и сходные с ним формы дали основание нескольким производным формам. Представители одной из них продолжили основную приспособительную линию лошадей - опору на центральный палец, но сохранили еще по два боковых пальца. Их адаптация шла по линии увеличения массы тела и толщины кожи, которая защищала тело одним или двумя кератиновыми образованиями на лицевой части черепа. Это были носороги (боковые пальцы им нужны для сохранения равновесия такой огромной массы). Некоторые носороги сохранились до наших дней в более или менее первоначальном облике, другие - высоко специализировались в собственном направлении и ушли от первоначального типа.
Вторая приспособительная линия родственна носорогам, хотя и древнее их. У тапиров боковые пальцы еще больше, каждая передняя нога имеет по четыре, а задние, как и у носорога, - по три пальца. Дополнительный палец на передней ноге расположен снаружи и предназначен для раскапывания почвы, разрывания корней и т. д. Масса тела также опирается на центральный палец. Эти живые ископаемые сохранились на протяжении 50 миллионов лет, и сегодня их можно встретить в тропических лесах Юго-Восточной Азии и Латинской Америки.
Представители третьей линии пошли в своей адаптации еще дальше, чем носороги. Они достигли гигантских размеров (со слона) и имели на морде раздвоенный рог. Это линия бронтотериев, которая расцвела в олигоцене и вымерла 30 миллионов лет назад.
Четвертая линия умеренно совместила в себе приспособления всех предыдущих, но несколько "качнулась" в свою сторону: развились огромные когти на ногах. Напоминали эти животные современных тапиров, значительно превышая их размерами. Жили халикотерии в лесу и саванне, питались корнями, клубнями и ветвями деревьев. Они претерпели менее эффектную эволюцию, чем бронтотерии, поэтому сохранились почти до наших дней - до среднего плейстоцена.
И, наконец, пятая линия прогрессировала не так ярко, как предыдущие, и дожила до наших дней, воплотившись в лошадь. К началу олигоцена в результате эволюции эогиппусов в Северной Америке появляются орогиппус, а затем мезогиппус и миогиппус. Эти животные были несколько крупнее родительской формы, величиной с овцу. Голова у них увеличилась, на ногах было по три пальца, хотя рудимент пястной кости четвертого пальца еще сохранялся. Кости пясти и плюсны, соответствующие функционирующим пальцам, стали значительно длиннее, чем у эоценовых форм. Все пальцы опирались на землю, хотя средний был крупнее и длиннее остальных. Очень изменилась зубная система: коронка коренных оставалась еще низкой, но предкоренные зубы по характеру жевательной поверхности уподобились коренным. И те, и другие зубы стали лофодонтными (вооруженными бугорками-шипами). Усиление зубной системы сказалось на лицевой части черепа. Она значительно вытянулась, глазница передвинулась к мозговой части черепа, увеличился объем мозга. Характер приспособлений этой формы позволяет сделать вывод о том, что начался постепенный выход этих животных из леса на опушки и в саванны. В их рационе стали преобладать не листья, а травы, на пережевывание которых приходилось тратить гораздо больше усилий.
Миогиппусы гораздо резвее стали спасаться от врагов на открытом пространстве.
В нижнем миоцене Северной Америки широко распространились парагиппусы. У них строение коренных зубов усложнилось и в углублениях между гребнями зубов появился цемент. Лицевой отдел черепа несколько удлинился, глазница стала частично замыкаться сзади отростком лобной кости. Боковые пальцы на ногах еще больше укоротились. От этого рода произошел меригиппус, у которого конечности стали длиннее, а боковые пальцы короче. Коронки коренных зубов приобрели призматическую форму, хорошо стал выражен цемент - признак, указывающий на то, что животные становятся травоядными.
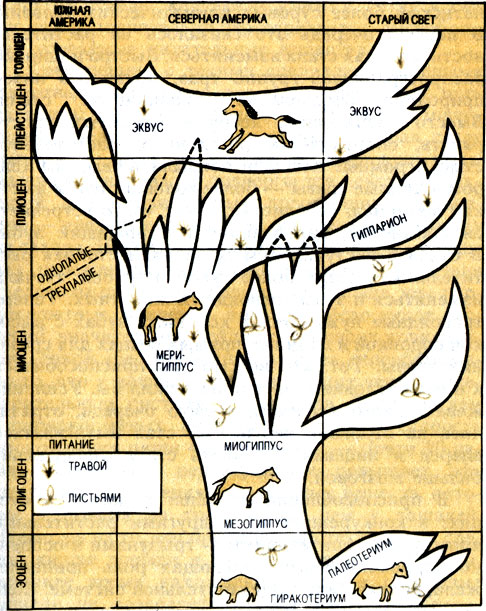
Эволюция рода лошадей (по Симпсону, 1951)
В миоцене произошло событие, значительно повлиявшее на развитие форм лошадиных. Климат на планете стал более континентальным, зима и лето обозначились гораздо четче, а тропики отступили к экватору. Та зона планеты, которую сегодня занимают леса и степи, оказалась под натиском более суровых климатических условий. В первую очередь это сказалось на растительности, которая стала изменяться. Быстро и широко распространилась группа трав, появилось новое природное образование - травянистые степи. Вместо тропических лесов и открытых пространств, спорадически заселенных растительностью, огромные площади покрылись зеленым ковром. Степные травы - более волокнистый и жесткий материал, чем листья, в них часто встречается кремнезем, царапающий и стирающий зубы. В связи с наступающим осушением степная растительность стала более сухой и грубой. Начали изменяться и зубы травоядных животных. Теперь травоядные нуждались в коренных зубах с высокой коронкой и длинных прямых резцах для срезания травы. Тот, кто не смог бы приспособиться к новым условиям, просто не выжил бы. Усиление жевательного аппарата, в свою очередь, отразилось на строении черепа: он стал вытягиваться вперед и лицевая его часть стала значительно больше мозговой.
В приспособлении лошадей к жизни в степях, в конкуренции их с другими растительноядными млекопитающими - грызунами и особенно парнокопытными решающая роль принадлежала, конечно же, пищеварительной системе. Если у парнокопытных (бычьих, антилоп, оленей, козлов, баранов) желудок четырехкамерный, то у лошадиных он однокамерный. Парнокопытное животное, наполнив на пастбище один из отделов желудка - рубец - зеленой массой, в укромном уголке пережевывает ее еще раз. К тому же в рубце на пищевую массу действуют многочисленные микроорганизмы, которые разлагают клетчатку. После второго пережевывания пища поступает в другие отделы желудка - книжку, сетку и сычуг, где дополнительно переваривается, высвобождая усвояемые питательные вещества.
Конкуренция с такими специализированными животными заставила лошадей найти свой путь развития: при однокамерном желудке у них значительно увеличилась слепая кишка, которая стала выполнять функцию рубца у парнокопытных. Здесь пищевая масса подвергается действию бактерий и многочисленных простейших организмов - инфузорий, которые разлагают клетчатку на сахара. Поскольку лошадям не нужно было еще раз пережевывать съеденную пищу, они стали съедать на пастбище травы вдвое больше, чем парнокопытные, причем - большее число видов растений, значительно обойдя в этом своих конкурентов. Такой "бесперебойной работе" способствовало то, что у лошадей пища проходит через пищеварительную систему в два раза быстрее, чем у парнокопытных. Например, у лошади скорость прохождения пищи 48, а у бычьих - почти 90 часов. Длительное переваривание повышает эффективность усвоения питательных веществ, которое у бычьих на треть выше, чем у лошадей. Несмотря на то, что процессы, протекающие в слепой кишке лошади и желудке жвачных, сходны, меньшее время нахождения пищи в организме лошади не позволяет стенкам кишечника переварить и поглотить максимальное количество питательных веществ. Поэтому остатки массы, выходящие из организма лошади наружу, остаются полупереваренными. Такой на первый взгляд незначительный факт имел громадное значение для формирования... самой степи.
Получив специфические приспособления, позволяющие выжить в сухих и бесплодных местообитаниях, на кормах, жестких и безвкусных для других копытных, лошадиные стали кочевыми животными. Избегая прямой конкуренции с парнокопытными, они, стравив какое-либо пастбище, перекочевывали на другое, затем на третье и т. д., забирая, таким образом, благополучно "свою долю".
Тысячекилометровые просторы степей манили травоядных копытных растительными богатствами. Для их освоения требовались более длинные и быстрые ноги. Поэтому наряду с жевательной и пищеварительной системой изменилась и вся конструкция организма лошадиных. Ноги стали длиннее, и еще больше дифференцировалось развитие центрального и боковых пальцев. Глазницы расположились так, чтобы в поле зрения входило все окружающее пространство. Лицевая часть относительно всего черепа укрупнилась, усложнились и увеличились зубы.
От меригиппусов произошла новая форма лошадиных - гиппарионы. Они были трехпалыми, но центральный палец настолько удлинился, что животное полностью могло опираться на него. Боковые же пальцы едва касались земли и прилегали к центральным. Все это было соединено со стопой и пястью мощной системой связок, позволяющей пальцам двигаться только вперед и назад, но не в стороны. Зубы гиппарионов укрупнились, бугорки на них объединились в длинные складки с цементом между ними. Лицевая часть черепа удлинилась, значительно усилилась лицевая мускулатура, особенно жевательная. Эта форма была явно похожа на настоящую лошадь, и она же оказалась одной из самых многочисленных и удачливых.
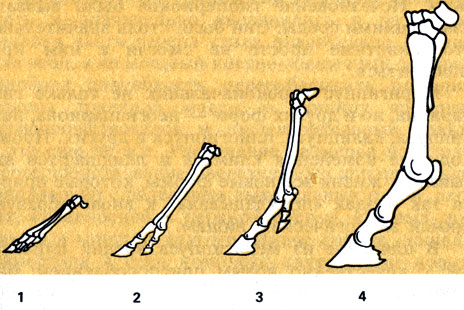
Эволюция конечностей лошадей: 1 - гиракотериума; 2 - мезогиппуса; 3 - меригиппуса; 4 - современной лошади
Время существования рода гиппарионов составляет около 11 миллионов лет, и только для Евразии уже описано 76 (!) видов и подвидов. Около 12 и повторно около 6 миллионов лет назад гиппарионы через Берингию как по мосту проникли из Северной Америки в Евразию, широко распространились по ее степным просторам и перебрались в Африку. Практически неизменными они дожили до среднего плейстоцена, и на африканском континенте бок о бок существовали с зебрами. Причина жизнестойкости гиппарионов заключена в их высокой приспособительной способности, благодаря чему они сумели заселить разнообразные экологические зоны - степи, саванны, поймы, редколесья. Продолжающиеся потепление и иссушение климата создавало новые ландшафты, которые гиппарионы осваивали и заселяли. Естественный отбор позволил "вылепить" форму, наилучшим образом приспособленную к новым условиям. Исчезновение гиппарионов было вызвано изменениями среды. Они были столь значительны, что животные просто не смогли к ним приспособиться.
Меригиппус - родоначальник не только гиппариона, но и других форм - неогиппариона, нан-ниппуса, калиппуса, плиогиппуса и других. Происходящие изменения климата и ландшафтов вызывали к жизни все новые формы, которые природа так и эдак "прикладывала" к вновь образующимся экологическим нишам.
В плиоцене из меригиппуса возник плиогиппус с качественно новым приспособлением: был однопалым. Он прочно обосновался на жительство в североамериканских прериях, и в конце эпохи из этой формы произошел род лошадей.
Появление собственно лошадей было скачком эволюции непарнопалых. Их отличия от более древних родительских форм на первый взгляд незначительны. Но обладая качественно новым уровнем приспособляемости, они в совокупности позволяют форме первенствовать в своей экологической зоне. Лошади почти полностью утратили боковые пальцы, от них остались лишь небольшие рудименты - грифельные косточки. Стопа и вся нога стала теперь располагаться перпендикулярно поверхности почвы, все кости конечности удлинились. На оставшийся средний палец приходилась теперь вся нагрузка тела, поэтому он стал крупнее и длиннее, а связочный аппарат, соединяющий его с метаподиями (пястью и плюсной), позволил ему теперь вращаться только в одной плоскости - вперед и назад. Ноги у лошадей заканчиваются крупным копытным "башмаком" - своеобразным стабилизатором при соприкосновении конечности с почвой.
Коренные зубы приобрели сильную складчатость и значительно увеличились в размерах. Лицевая часть черепа стала значительно больше мозговой, а ее мощный костно-мышечный аппарат приобрел способность перерабатывать большие объемы травяной массы в единицу времени. Усовершенствовался и пищеварительный тракт. И это еще не все. Увеличение размеров ног, тела и совершенствование двигательного аппарата шло параллельно с развитием дыхательных путей, легких, кровеносной и нервной систем, кожных покровов и многого другого. Иначе говоря, лошадь совместила в себе комплекс качественно новых признаков, позволивший ей подняться на более высокую ступень лестницы приспособлений и стать составной частью равнинных ландшафтов.
В нижнем плейстоцене лошадиные широко расселились в Северной Америке, а затем проникли в Евразию. Некоторое время они существовали рядом с гиппарионами, но постепенно те стали вымирать. К концу плейстоцена в Америке вымерли все лошадиные, освободив жизненное пространство для других групп животных. Гонка за выживание продолжалась.
В среде современных палеонтологов и сегодня нет единства в вопросе происхождения под-родов лошадей, ослов и зебр. Господствует мнение, что все они произошли от предка, который уже был однопалым и имел сложные зубы с высокими коронками (гипсодонтные).
Как бы там ни было, а в плиоцене Северной Америки от разновидностей плезиппуса произошли две группы лошадиных. Одна из них крупнее, напоминает современную зебру Грэви и появилась 5 млн лет назад, а вторая, сходная с бурчелловой зеброй, сформировалась на 1,5 млн лет позже. Климат в то время в Северной Америке был мягкий, на месте сегодняшних прерий находились влажные саванны.
Около 3 млн лет назад зебры расселились и в Европе. Эти животные имели удлиненный череп с мощным зубным аппаратом и тонкой сложной эмалью на коренных зубах. Группа крупных зебр была представлена в Европе лошадью Стенона.
Начавшееся в плейстоцене похолодание климата и образование ледников привели к появлению в Европе и Азии массивных форм лошадей с еще более мощным зубным аппаратом, толстой кожей и густым шерстным покровом. А теплолюбивые зебры начали вымирать. Если бы в плейстоцене отдельные формы зебр не перебрались в Африку, мы с вами, видимо, так и не знали бы, что есть полосатые лошади. В благоприятном климате континента зебры сохранились почти неизменными, донеся до нас дуновение эпохи пещерных львов, махайродов, мастодонтов и эласмотериев.
Плиоценовые лошади Азии, обладая в общем всеми прогрессивными признаками их современников из Европы, тем не менее шагнули вперед. Строение зубов ископаемых азиатских лошадей (развитие внутренних складок) показывает, что среда заставила эти формы лошадей приобретать новые приспособительные черты. Возможно, азиатские лошади произошли от американских плиогиппусов с уже более развитой зубной системой, а возможно, что потомки плиогиппусов из Европы распространились в Азию, где и приобрели более прогрессивную зубную систему. Это близкие формы - сиваликская и санменская лошади. Они населяли сухие дерновинные степи Центральной Азии - регионы с устойчивым климатом. У них были прямая линия профиля черепа, длинные и тонкие конечности с узкими копытами. Впоследствии эти формы образовали группу полуослов и группу настоящих лошадей.
Эволюция настоящих лошадей
В древнейшем плейстоцене (1 млн лет назад) животный мир зоны современных степей от Причерноморья до Маньчжурии имел африканский облик и существовал в саванноподобных ландшафтах при умеренно теплом климате. Здесь обитали двурогие носороги, лошади Стенона, ослы, винторогие антилопы, кабаны, лоси, газели, верблюды, бизоны, различные олени. Наступившая перед этим ледниковая эра вызвала резкое похолодание, увлажнение, значительные геологические и ландшафтные перестройки. Часть тропической фауны в эту эпоху вымерла, часть отступила в более южные районы Средиземноморья и Передней Азии, а оставшиеся начали постепенно приспосабливаться к изменившейся среде.
В раннем плейстоцене (700-350 тыс. лет назад), во время миндельского оледенения, ландшафты средних широт Евразии были уже заняты лесостепями умеренного пояса. К этому времени полностью исчезли теплолюбивые антилопы, широко распространился бизон, появились холодостойкие формы лосей и носорогов. Изменения среды заставили изменяться и лошадей.
В Европе из наиболее крупной формы - лошади Стенона - появились гигантские лошади нескольких форм, получившие у палеонтологов наименования: мосбахская, тираспольская, лошадь Абели. Они и были собственно первыми настоящими лошадьми в Европе. Эти лошади утратили многие архаичные черты в строении черепа, зубов и конечностей, изменились общее сложение и пропорции тела. Они стали более тяжелыми, с массивными ногами, широкими копытами. Голова вытянулась, стала длинномордой и узкой. Зубы у них были мелкими, с несколько ослабленной складчатостью. Кожа стала толще, а волосяной покров гуще и грубее.
В это же время в зоне современной тундры - в Берингии, Якутии и на Таймыре - формировалась арктическая фауна копытных. Ландшафты были здесь представлены тундростепями, по долинам северных рек тянулись лиственные леса и кустарники. Арктическая фауна копытных формировалась в одно время с предками позднейших лошадей, овцебыков, северных оленей.
Средний плейстоцен был эпохой расцвета ранней мамонтовой фауны, что могло произойти только в условиях широкого развития степных и лесостепных ландшафтов. В это время по равнинам Восточной Европы и Сибири расселились крупные лошади, осел, огромный верблюд, гигантский и благородные олени. Наибольших размеров и численности достигли овцебык, длиннорогий бизон, первобытный тур. В Европе обитали разнообразные формы лошадей, близких как к лошади Стенона, так и к уже образованным из них новым, более прогрессивным формам. Дело в том, что резкие сдвиги в геологии и климате Европы вызвали появление и развитие в различных местах как новых форм лошадей, так и совершенствование в каком-то направлении старых, уже приспособившихся форм.
Здесь же, в среднем плейстоцене, в хозарскую эпоху существовали лошади уже совершенно современного облика, среднемордые и среднелобые, с массивными конечностями и зубами с утолщенным эмалевым слоем. Обозначились и географические разновидности лошадей.
Форма, обитающая в лесостепях и лесах севера Восточной Европы, имела более длинномордый и узкий череп с относительно мелкими коренными зубами. Эти лошади жили рядом с лосями, бобрами и медведями. Более южная форма, поселившаяся вместе с верблюдами, сайгаками и ослами, была значительно крупнее предыдущей, короткомордой, широкоголовой, с более крупными зубами. Несомненно, она более прогрессивна. Основным фактором, ускоряющим развитие ее приспособительских качеств, был, конечно же, степной ландшафт.
В верхнем плейстоцене Европы эпохи похолодания и потепления чередовались. В эпохи похолодания появились крупные, массивные формы лошадей, так называемые широкопалые лошади. В эпохи потепления их место занимали более мелкие, широколобые, короткомордые и тонконогие формы, обитавшие на юге Западной Европы, в Закавказье и Юго-Западной Азии.
Сегодня сложно сказать, что происходило с приходом ледников. Уходили на юг одни формы лошадей, а им на смену приходили с севера другие, более холодостойкие и более приспособленные? Или теплолюбивые формы постепенно, на протяжении сотен поколений преобразовывались в холодостойкие? Оценивая систему поведения современных лошадей, преодолевающих в миграциях сотни километров, нужно отдать предпочтение первой гипотезе. Но тогда откуда появились более приспособленные, холодостойкие формы?
В позднем плейстоцене (70-60 тыс. лет назад) Евразии, в эпохи последних оледенений, достигла своего расцвета поздняя мамонтовая фауна: шерстистый носорог, мамонт, три-четыре вида широкопалых лошадей, плейстоценовый осел. В Южной Сибири и Забайкалье широко распространились по долинам рек марал и гигантский олень. Все равнины от Атлантического побережья Европы до Аляски заселили тысячные стада бизонов, лошадей, сайгаков, куланов, овцебыков, северных оленей, которые паслись вместе с мамонтами. Все они отлично освоились в сухом холодном климате и ландшафтах тундрово-степного типа.
В послеледниковую эпоху (12-10 тыс. лет назад) в Евразию пришло великое потепление. Климат увлажнился, и лёссовые тундростепи стали постепенно превращаться в заозерные и замшелые тундры, а равнины - в долинные и горные леса. В этот период происходили резкое сокращение ареалов и частичное вымирание степных видов копытных: гигантского оленя, первобытного бизона, тура, байкальского яка, овцебыка. Более половины видов копытных в Евразии вымерли, а остальные - лошади, бизоны, северный олень, козлы и другие стали приспосабливаться к новым условиям. Им предстояло жить в лесах, тундрах и частично в горах. В голоцене образовались некоторые лесные формы лошадей, дожившие почти до настоящего времени в Прибалтике, Прибайкалье и на Аляске.
В позднем палеолите (9,5 тыс. лет назад) в ареалах восточноевропейских (мезенских) лошадей, населявших степи Южной и Средней России, появились и широко распространились формы, которые были предшественниками неолитических лошадей Восточной Европы. Именно они и были одомашнены степными кочевниками.
Неполнота палеонтологической летописи не позволяет в ряде случаев судить определенно о тех или иных изменениях в эволюции рода лошадей. Есть данные, что тяжелые лошади отступили в палеолите в лесную зону, а им на смену из Юго-Западной Азии пришли более легкие чисто степные формы, часть которых и была одомашнена. Другие оставались в диком состоянии вплоть до конца XIX в. в таврических степях и известны под названием "восточноевропейский тарпан".
История рода лошадей в Азии значительно отличается от таковой в другой части света. Основное отличие обусловлено своеобразными палеоландшафтными условиями послетретичного периода. В силу обособленности и огромной протяженности в Азии господствует континентальный устойчивый климат, поэтому здесь - царство архаичных форм живых существ, сохранившихся со времен последних резких изменений на материке и отставших в своем развитии от европейских. Наличие в Азии многих экологических зон (тундры, лесов, гор, степей, пустынь) вызвало к жизни развитие нескольких групп фаун: тундрово-степной, лесной, плоскогорной, полупустынной. К тому же в восточных и западных, северных и южных областях материка формировалась своя специфическая фауна. Открытые тундрово-степные ландшафты с достаточно твердыми грунтами на севере, лесные, лугово-степные и лесостепные в центральной части и пустынно-плоскогорные на юге оказались благоприятными для обитания лошадей. Соответственно этим биотопам образовывались и различные формы лошадей На севере возникли тундровые формы, населявшие в плейстоцене и позже тундру от Аляски до Таймыра, - колымская, ленская разновидности якутских плейстоценовых, дикие лошади Аляски. Все они были небольшие, приземистые, крепкие, с большой грубой головой и светлой мохнатой шерстью.
Южнее, на границе лесов и степей, сформировались южносибирские и бурят-монгольские лошади, а также форма, которая со временем, перейдя с человеком севернее, в Якутию, стала современной якутской лошадью. Это тоже приземистые лошади, светлой и серой (мышастой) окраски. На юге ареала, в предгорьях Тянь-Шаня и Монгольского Алтая, в Джунгарии, на равнинах Гоби и в Среднем Китае сформировался степной тип лошадей, давший несколько разновидностей монгольских и китайских лошадей, а возможно, и современной азиатской дикой лошади. Этот тип образовался из форм группы санменской лошади.
Последняя из лошадей, известная как лошадь Пржевальского, отличается от других близких форм длинномордостью, крупными коренными зубами, менее складчатой и более толстой эмалью коренных зубов, очень тонкими фалангами пальцев. Лошадь Пржевальского отличается от всех других современных и плейстоценовых лошадей Европы и Северной Азии. Она не могла сложиться нигде, кроме как в Центральной Азии, и несет на себе печать более ранних этапов развития Земли, видимо, позднего плейстоцена. Устойчивый климат Центральной Азии способствовал сохранению этого типа до настоящего времени почти в неизменном состоянии с тех времен, когда в Азии впервые появились первые настоящие (кабаллоидные) лошади.
Немного о систематике
Отряд непарнокопытных (Perissodactyla Owen, 1845) вписал в летопись планеты свою собственную историю с раннего эоцена по сегодняшний день. Пора расцвета представителей отряда, к сожалению, давно прошла. И неудивительно, ведь позади более 55 млн лет их истории. Это реликтовые млекопитающие, в живых остались лишь несколько видов с малой численностью.
Отряд состоит из двух подотрядов - Hippmorpha и Ceratomorpha. В первом три семейства, включающих более 75 родов, из которых до современности дожил только один род (Equus), остальные все вымерли.
Из второго подотряда, состоящего из семи семейств, сохранились только два рода - тапиры и носороги, причем в первом - только один вид из пяти, а во втором - четыре из 35! Сколько же всего было вымерших родов, видов, подвидов? Это неизвестно.
Семейство лошадиные (Equidae Gray, 1821) состоит из нескольких вымерших подсемейств - Хиракотерины, Анхитерины, Мерихиппины и одного современного - Эквины. История последнего начиналась в позднем плиоцене и разворачивалась на территории Америки, Евразии и Африки. До настоящего времени сохранился только род лошади (Equus Linnaeus, 1758), а вымершие представляют собой переходные формы от гиппарионов к лошадям. Этот род включал в себя около 13 подродов, но плиогиппусы, плезиппусы, аллогипусы и другие давно вымерли, и теперь в нем осталось лишь пять подродов: Dolichohippus Oustalet, 1882 - зебра Греви; Hippotigris Smith, 1847 - все мелкие зебры; Asinus Frisch, 1775 - настоящие ослы: Hemionus Stehlin et Graziosi, 1935 - полу ослы, или куланы; Equus Linnaeus, 1758 - настоящие лошади.

Н.М.Пржевальский (1839 -1889) - генерал-майор, знаменитый русский путешественник, первооткрыватель многих областей Центральной Азии и Сибири. Открыл для науки ряд неизвестных видов животных, в том числе дикого верблюда и лошадь Пржевальского
Последний подрод состоит из 7-10 видов собственно настоящих лошадей, история которых начинается с раннего плейстоцена и продолжается по сегодняшний день. Все дикоживущие формы, кроме лошади Пржевальского, вымерли, известны они только по ископаемым останкам.
Раннеплейстоценовая лошадь когда-то населяла территорию Молдавии и Румынии. Мосбахская лошадь - крупная плейстоценовая форма, обитавшая в Южной и Центральной Европе. Очень крупная европейская лошадь Абели жила во времена миндельского оледенения.
Штейнхетская ледниковая лошадь в середине плейстоцена населяла Европу, а широкопалая в среднем и позднем плейстоцене - Восточную Сибирь и европейскую часть СССР. В ту же эпоху жили германская лошадь - на территории Центральной Европы, уральская - севернее Каспийского моря и ленская - в Сибири, от Чукотки до Таймыра. Тарпан восточноевропейский населял степи Восточной Европы вплоть до конца прошлого века (последний тарпан погиб в 1879 г. в Агайманском поду, севернее Аскании-Нова).

И.С.Поляков (1845- 1887) - член Императорского географического общества, ученый-хранитель отделения млекопитающих Зоологического музея, магистр зоологии, революционер, автор первоописания лошади Пржевальского (1881 г.) по материалам, доставленным Н. М. Пржевальским
За последние 100 лет, с момента описания дикой азиатской лошади и по сей день, не утихают споры ученых о ее месте в систематике. Первоописание этой лошади (Eguus (Eguus) przewalskii Poljakov) было сделано в 1881 году ученым - хранителем отделения млекопитающих Императорского зоологического музея магистром зоологии И. С. Поляковым (1845-1887) по шкуре и черепу, доставленным Н. М. Пржевальским из Центральной Азии. И. С. Поляков определил, что это особый вид, который не может быть тарпаном (восточноевропейским - В. К.) и по своей внешности занимает середину между дикими азиатскими ослами и нашей домашней лошадью.
У И. С. Полякова сразу же нашлись противники, в основном в Западной Европе. Европейские ученые Матчи в 1903 г. и Хильцхеймер в 1909 г. описывают уже три вида лошади Пржевальского (Е. hagenbecki, Е. equiferus tipicus и Е. przewalskii). Немецкие систематики Лидеккер и Шварц отказывают дикой лошади в праве именоваться "вид" и превращают ее в подвид домашней лошади (Е. caballus przewalskii). Наш отечественный зоолог В. Г. Гептнер считал, что лошадь Пржевальского и степной тарпан представляют собой не более чем географические, правда резкие, расы дикого вида и под общим названием Eguus przewalskii объединил в качестве подвидов всех голоценовых лошадей, включая восточноевропейского и лесного тарпанов, домашнюю лошадь, лошадь Пржевальского и прочих диких настоящих лошадей. Его поддержали некоторые зарубежные авторы, объединившие диких азиатских лошадей с домашними и дикими европейскими. Огромный вклад в дело сохранения видовой самостоятельности лошади Пржевальского внесла известный советский палеонтолог В. И. Громова. Изучив ископаемые костные останки древнейших лошадей и соотнеся их с костями современных и лошадей Пржевальского, она пришла к выводу, что лошадь Пржевальского отличается от позднеплейстоценовых и современных лошадей рядом признаков (длинномордость, крупнозубость, тонконогость), характеризующих ее как более ранний, примитивный тип, самостоятельно развивавшийся с нижнего плейстоцена, что позволяет выделить его в самостоятельный вид, а предположительным предком лошади Пржевальского является санменская лошадь, на что указывает ряд общих признаков.
Сегодня видовая самостоятельность лошади Пржевальского подкреплена исследованиями ее морфологии, физиологии, поведения и т. д.
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://konevodstvo.su/ 'Konevodstvo.su: Коневодство и коннозаводство'